氧化亚氮(N₂O)是一种强效且长寿命的温室气体,在百年时间尺度上,其增温潜势约为CO₂的近300倍。尽管科学界对全球氮循环进行了深入研究,但N₂O的来源归因仍存在较大不确定性。这种不确定性在一定程度上源于我们对“硝化菌反硝化” (nitrifier denitrification) 过程的认知有限。
氨氧化细菌(AOB)是硝化微生物的重要类群,通常被认为是专性好氧微生物,依靠氧气将氨氧化为亚硝酸盐来获取能量。然而,一个长期困扰学术界的现象是:在稻田、湖泊沉积物以及海洋最小含氧带(OMZs)等缺氧甚至厌氧生境中,依然能观测到大量AOB的活跃存在。前期一些工作,包括团队新近的研究(Nature Geoscience, 2024 17:1254-1260),表明在这些氧受限环境中,AOB的生存可能与硝化菌反硝化过程相关;这一过程也被推测是N₂O排放的重要来源之一。但是长期以来,关于AOB在缺氧条件下通过该途径究竟只是“解毒”,还是也能从这一过程获得能量以维持生长与代谢,一直未有定论。
2026年1月20日,浙江大学生命科学学院程磊教授-肖璟研究员团队在国际知名学术期刊《Soil Biology & Biochemistry》在线发表题为“How nitrifiers denitrify?”的观点分析文章,提出一个关于硝化菌反硝化的概念性框架,为理解AOB在缺氧环境下通过硝化菌反硝化作用维持生存提供了新思路。

团队系统梳理了近期关于亚硝酸盐还原酶和一氧化氮还原酶的酶学和热力学研究进展。文章指出,传统关注的含铜型亚硝酸盐还原酶(NirK)以及细胞色素c依赖型一氧化氮还原酶(cNOR),缺乏与跨膜“能量获取”相耦合的结构基础;同时,NirK的表达主要受亚硝酸盐积累影响而非低氧直接调控,因此更可能主要承担降低亚硝酸盐及相关中间产物毒性的作用。相比之下,AOB体内还广泛存在一种含血红素-铜中心的一氧化氮还原酶(sNOR)。根据理论推测,sNOR活性位点具有保守的质子通道结构:当AOB利用sNOR将NO还原时,可能同时跨膜转运质子并形成质子驱动力(PMF),从而在以氢气、丙酮酸等非氨底物作为电子供体的厌氧呼吸中获得可用能量(图一)。
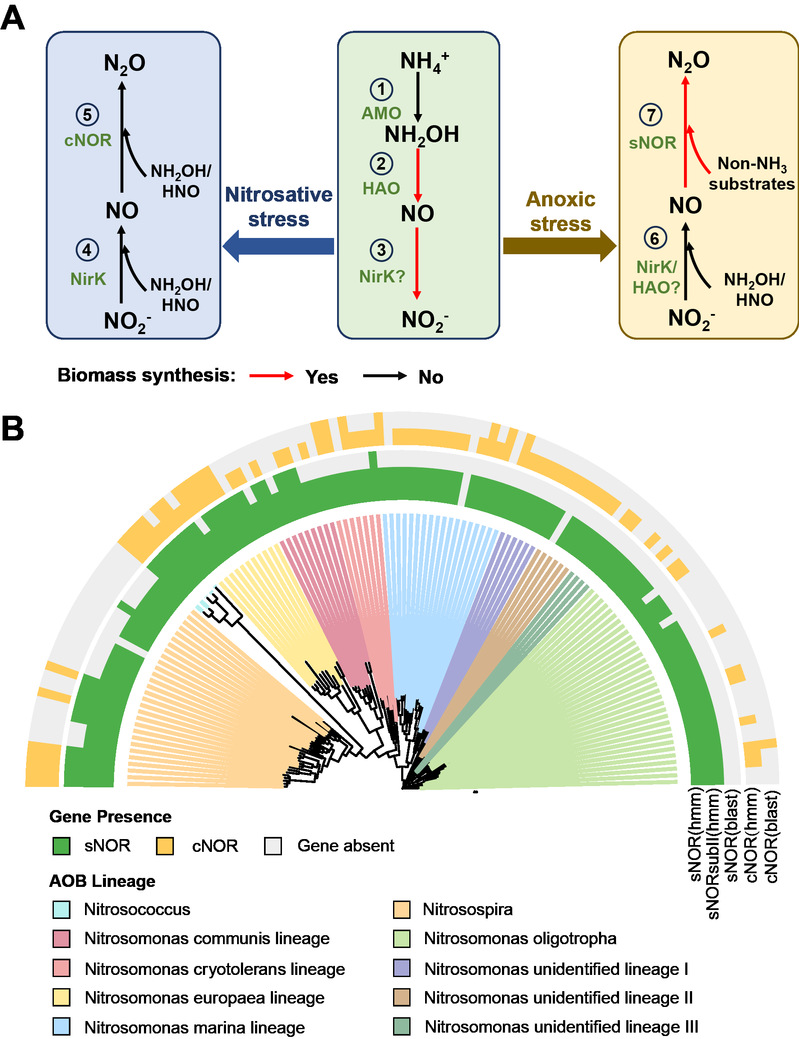
图一:AOB进行硝化菌反硝化的生理基础与系统发育分布
通过对全球氨氧化细菌基因组数据库的系统发育分析,团队进一步发现,sNOR基因存在于近96%的已测序AOB基因组中,涵盖了从陆地到海洋的广泛谱系。相比之下,cNOR仅存在于约56%的AOB中,并在部分与缺氧生境相关的谱系中缺失。这一结果表明,利用sNOR介导反硝化作用来获取能量,可能是AOB应对缺氧胁迫的一种普遍且核心的生存策略。
文章进一步指出,随着气候变化导致的全球水体和低地生态系统“脱氧” (deoxygenation)趋势加剧,具备sNOR的AOB可能会在未来的缺氧环境中占据竞争优势。这将导致硝化菌反硝化过程成为全球N₂O排放的一个重要且可能持续增长的来源,尤其是在受气候变化影响显著的水生和低地生态系统中。文章提出,未来的气候模型亟需重新评估硝化菌反硝化的贡献,以更准确地预测全球N₂O排放趋势。
浙江大学生命科学学院张凯杭博士为本文第一作者,程磊教授为通讯作者。特别感谢加拿大阿尔伯塔大学Lisa Y. Stein教授、课题组徐陈超博士和肖璟研究员为本文提供的建设性意见。该研究得到了国家自然科学基金及浙江省自然科学基金的资助。
原文链接: https://doi.org/10.1016/j.soilbio.2026.110098